Da Capo
 by Grady Towers
by Grady Towers
I wanted to know if it was possible to cross a human being with a chimpanzee and obtain a viable offspring. I knew of course that man has 46 chromosomes and the chimpanzee, like the other great apes, has 48. But I also knew that the horse has 64 chromosomes and the ass has 62 and they can still be crossed to produce a mule with 63. That implied that a difference in chromosome compliment might not be the barrier to breeding that it first appeared to be. I also knew that the DNA sequences of man and chimpanzee were identical at 99 out of 100 base pairs. The possibilities for a viable cross, therefore, looked reasonably good - good enough at any rate to justify spending some time in a library researching the matter.
At first I considered trying to find out if anyone had actually carried out such an experiment. Mankind's sexual propensities being what they are, I wouldn't have been surprised; there's a good reason for syphilis being named for a mythological shepherd. But I finally discarded this approach as unproductive; I found myself chasing rumors of Bigfoot and yeties, rather than securing information I could trust. So I eventually turned to experimental genetics for my data, and there I found the answer to my question. The answer came from an article titled "The Striking Resemblence of High-Resolution G-Banded Chromosomes of Man and Chimpanzee," written by Jorge J. Yunis, Jeffrey R. Sawyer and Kelly Dunham [Science, Vol 208, 6 June 1980, pp. 1145-8]. These investigators applied a new staining agent called giemsa to the chromosome compliments of man and chimpanzee and made a detailed comparison of their banding patterns. Their new stain was able to resolve more than a thousand bands in the chromosomes of each species, revealing a similarity so close that they found it difficult to account for the phenotypic differences. As part of their article, they provided a diagram of the chromosome comparisons, showing not only an astonishing similarity, but a number of interesting differences as well. Among these differences was the revelation of nine pericentric inversions. This observation provided the answer to my question. Chromosome inversions are known to result in semi-sterility when crosses are made to individuals without the inversion. Since there are nine of these, and since a cross with only one inversion results in semi-sterility, the answer must be: No, it's not possible to cross a human being with a chimpanzee and obtain a viable offspring.
Sometimes, however, when one is looking for an answer to a trivial question, one stumbles across the answer to a much more important one. That's what happened in this case. The diagram given in the article clearly reveals the exact genetic mechanism responsible for the evolution of genus Homo, and strongly suggests that this did not take place over hundreds of thousands of years, as is generally believed, but occurred within the span of only three generations.
The theory of evolution taught in school when I was an undergraduate anthropology major was called the Modern Synthesis, a term coined by Julian Huxley in 1942. It attempted to integrate the insights offered by Darwinian evolution with those of modern population biology and genetics. Essentially it said that point mutation within structural genes was the source of variability within species, that the emergence of a new species was the result of the accumulation of many mutations, and that the pace of evolution was slow. Moreover, it said that the direction of evolutionary change was the result of natural selection working small variations. The shape of an organism, in other words, was the result of its adaptation to a specific environment. In effect, the theory said that the origin of a new species (macroevolution) was due to exactly the same causes as changes within a species (microevolution). But above all, it said that change was gradual.
As time went by, however, geologists and paleontologists began to find the theory unsatisfactory. No one doubted that the overall record showed a steady increase in the diversity and complexity of species, but it was becoming embarrassingly obvious that the fossil record did not show a smooth transition from one form to another. Instead, the record that species typically remained unchanged for millions of years, and then were abruptly replaced by a new species that were substantially different in form though clearly related. Evolution apparently worked in a jerky fashion, rather than in the smooth, gradual manner postulated by the Modern Synthesis.
The new view of evolution as characteristically jerky, now called punctuated equilibrium, was given its present form by Steven Jay Gould of Harvard and Niles Elderidge of the American Museum of Natural History in New York. They conceded that mutation and adaptation to specific environments, as postulated in the Modern Synthesis, applied to macroevolution as well as microevolution, but they also believed that another factor was at work in species formation. Among the most likely candidates for this factor was one called chromosome speciation. This theory, as proposed by Guy Bush of the University of Texas and Alan Wilson of Berkeley, says that a new species arises as a result of a re-arrangement of chromosome structure and not as a result of mutation. This proposal is a sound one. The connection between chromosome number and speciation is one kind of re-arrangement that is well known among plant breeders. They've known for a long time that by simply increasing the number of chromosomes typical of a species - a phenomenon called polyploidy - that it was possible to obtain a new variety with different characteristics. Many commercial plants, it turns out, were created in exactly this way; common bread wheat, for example, is a hexaploid, having three times as many chromosomes as its parent species, and some strawberries are octoploid, with four times the expected number.
But while polyploidy is fairly common among plant species, it's rarely found among animals, being observed mostly among those that reproduce asexually. Instead, the chromosome re-arrangement most often exhibited in animal species is translocation. This is when non-homologous chromosomes break and exchange parts: one of the two chromosomes in pair A exchange a part with one of the two chromosomes in pair B. If the break in the respective chromosomes occurs near their ends and the long parts are joined together, the short segments sometimes contain so little genetic information that they may be lost. This gives the appearance of two chromosomes having been fused together. This is what happened in our own ancestry, and is the mechanism responsible for the origin of man.
I have reproduced a small part of the diagram given in the article, showing the human second chromosome on the top, and two chimpanzee chromosomes on the bottom. It is patently obvious that the human second chromosome was created by translocation, or a fusion if you prefer, of two chimpanzee chromosomes. Or to be more precise, that the human second chromosome was created by a translocation of two chromosomes in an animal that was ancestor to both man and chimpanzee.
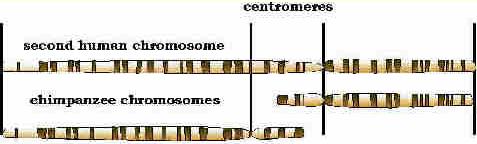
The sequence of events probably took place something like this. About five million years ago a translocation like that described above occurred in a pithecine male who was the controller of a harem of females. Rather than having 48 chromosomes, which was normal for his species, he had 47. When he mated with members of his harem, who possessed the usual number of chromosomes, half of his offspring would have had 48 chromosomes and half would have had 47. If some of those with 47 chromosomes mated among themselves, or were back-bred to their father, one quarter of their offspring would have had 48 chromosomes, one half would have had 47, and one quarter would have had 46. Those with 46 were the prototype of the new genus Homo. But at this stage they were not yet a new species. At most they can be thought of as a new chromosomal race., probably with great phenotypic difference from their fellows, but still not yet a new species. That had to wait for the appearance of one of the chromosome inversions discussed above. This inversion also probably occurred in a male with a harem and was transmitted in much the same way as the translocation. In this case, however, crosses between individuals with the inversion and those without produced only a few offspring, while matings between inverted chromosomes continued to be fertile, as did those without the inversion. This was the first step in breeding isolation. Suddenly, almost overnight, a new species came into existence.
I'm personally convinced that something like the scenario just presented really did take place. The exact details are almost certainly wrong, but the essentials ought to be correct. Still, it would be nice to have some experimental confirmation of the theory. Suppose we were to cut the human second chromosome in egg and sperm at exactly the same place it was originally fused together using recombinant DNA techniques. Could we back-breed man's pithecine ancestor? Could we recover "Lucy," the first member of our genus?
When this idea first occurred to me, I had a wonderful time working out the social and legal implications that would result if such an experiment could be carried out successfully. Unfortunately, the experiment won't work. Cutting the chromosome at the exact spot necessary is fairly simple in principle: one merely tailors the appropriate endonuclease for the point at which the cut is to be made. But the problem that can't be overcome is that only one of the two chromosome fragments would have a centromere, the indented part in the diagram. This is where the spindle fibers attach during cell reproduction, and without a centromere for every chromosome, the reproductive process fails. The cell dies.
On the other hand, another equally exciting experiment might well be possible. Suppose we applied DNA techniques to the chromosomes of chimpanzees. Suppose we attempt to fuse the same two chromosomes in chimpanzees that resulted in the origin of our own genus. Could we expect to get the same profound physical and mental changes that occurred in our own ancestry? And what are these changes likely to be? We know from Jane Goodall's work that chimpanzees are already tool users, and although scholars are bitterly divided on the subject, some believe chimps already have a rudimentary command of language. Furthermore, Arthur Jensen claims that the very brightest chimpanzees have the mental ability equivalent to that of the average nine or ten year old human [Bias in Mental Testing, p. 182]. Could an experiment like the one proposed tilt chimpanzees across the threshold into full intelligence?
Aside from possible mental changes that might result, we could also expect two important physical changes. One is an increase in neotony. Neotony means that the individual retains infant-like characteristics throughout its life span. When a human infant is compared to a chimpanzee or gorilla infant, their appearance turns out to be amazingly alike. But as each species matures, chimps and gorillas change greatly, whereas man continues to resemble his infant self.
The second physical change is likely to be more upright stature with the long human leg and striding walk. Much is made of man's opposing thumb, his stereoscopic color vision, his capacity for language, and so on; but his evolutionary success has been as much the result of his striding walk as all the more salient characteristics. If we were as short legged as chimps and gorillas - still knuckle-walkers - we would be confined to the continent of Africa. As it is, we spread over the entire face of the earth in a remarkably short time. Our chimpanzee protégés might be physically blessed in a similar way.
Could present DNA techniques really accomplish the fusion of two chromosomes? I admit that the outcome of such an experiment is far more problematical than simply cutting a chromosome in the right place. But if it is presently impossible, it soon won't be. A new engineering discipline called nano-technology will make the process child's play. The new technology is concerned with building microscopic machines and micromanipulators. Some of those working in the field believe that one day chemists may be able to physically manipulate individual atoms, to assemble and disassemble molecules as though they were tinker toys. If so, then the fusion of two chromosomes will be simple in comparison.
Very well, then, suppose the techniques work. Suppose that the experiment is tried and we do indeed obtain a viable, intelligent variety of chimp. Then what happens? Then the investigator applies for a patent. According to federal law, a new variety of life can be patented. Obviously, the law was intended to cover microbes that manufacture useful chemicals, or clean up oil spills. Obviously, it was not intended to cover the origin of a new, possibly intelligent species. Obviously, the case would go to the Supreme Court. One wonders how that court will deal with the hoary philosophical question of, "What is Man?"
Should such an experiment be carried out? For decades now, astronomers have been searching the heavens for signs of intelligent life. I submit that the reason for that search can be used as validly to justify the experiment I've proposed. Moreover, because they are our distant kin we have far more to learn from an intelligent race of chimps than we could ever hope to learn from some alien species in the stars: insights into intelligence, language, social organization and so on. But most of all, I would want to know if they had souls. The only way I can see to answer that question is to teach them one or more of the great religions and then to ask them. No matter what they might say, the answer is bound to be fascinating. Then I would ask them if they thought we had souls, too.











No comments:
Post a Comment