Speculations on the Brains and Minds of Cetaceans
 by Huntley Ingalls
by Huntley Ingalls
"Genius in the sperm whale? Has the sperm whale ever written a book, spoken a speech? No, his great genius is declared in doing nothing in particular to prove it. It is moreover declared in his pyramidical silence. And this reminds me that had the great sperm whale been known to the Orient World, he would have been deified by their child-magician thoughts…
"Champollion deciphered the wrinkled granite hieroglyphics. But there is no Champollion to decipher the Egypt of every man's and every being's face….If then, Sir William Jones, who read in thirty languages, could not read the simplest peasant's face in its profounder and more subtle meanings, how may unlettered Ishmael hope to read the awful chaldee of the sperm whale's brow? I but put that brow before you. Read it if you can.
"My object here is simply to project the draught of a systematization of cetology….But it is a ponderous task"to grope down into the bottom of the sea after them; to have one's hands among the unspeakable foundations, ribs, and very pelvis of the world; this is a fearful thing. What am I that I should essay to hook the nose of this leviathan!" - Herman Melville
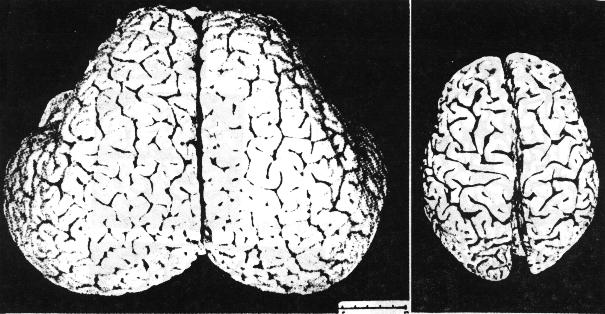
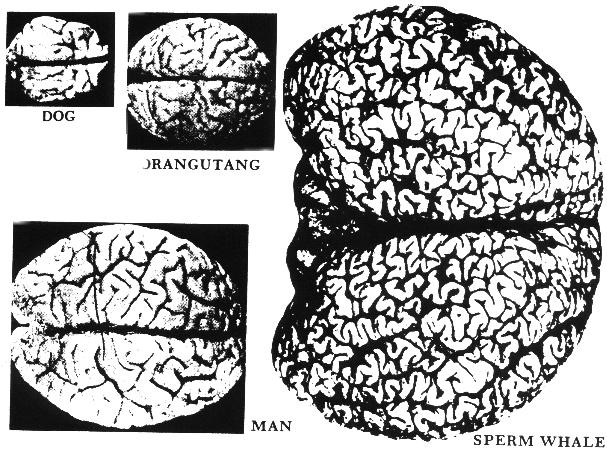
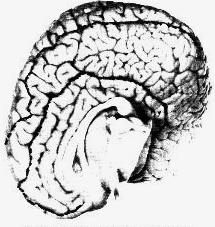
Anyone familiar with the brains of higher mammals is shocked when first confronted with the physical appearance of the cetacean brain. Its size, globular shape, and elaborate convolutions suggest what might be imagined as the brain of a superior being. The typical weight of a human brain is about three pounds, of a dolphin about 3.5, of a large baleen whale 13, and of a sperm whale 20. The convolutions of the cetacean brain are far more elaborate than those of the human brain. The density of neurons and the complexity of neural connections in the two brains are comparable. The thickness of the cetacean neocortex is less than that of the human, but its greater surface area supports, in some species, an equal or greater number of nerve cells. The early whales of Eocene times, about 50 million years ago, were fully aquatic but had small, primitive brains. Large-brained cetaceans appeared about 30 million years ago, and large-brained humanoids about 5 million years ago. The greater length of evolution for the cetacean brain is not proof of greater advancement, but it is a startling suggestion of refinement.
Brains are metabolically expensive and such huge neural systems cannot be mere embellishments. They are unnecessary for simply operating a large marine animal. The whale shark, which is sometimes 50 feet long, does well with its primitive fish brain. The enormous development of the cetacean brain has evolved through survival advantage or social selection and must support functions of great importance to whales. We will try here to imagine the possible nature of these functions. Most of the ideas presented are highly speculative, but at least some of them are testable in principle. Even wrong ideas are often valuable if they can stimulate further thought or suggest experiments and tests which sharpen old conceptual landscapes or illuminate new ones.
Whales have limited ability for physically manipulating objects. This alone would form their minds differently from human minds, because our concept of objects and physical space is greatly influenced by kinesthetic interaction with the environment. As far as can be determined, sound is the only familiar sensory mode in which they are extensively involved. This is a very effective sense mode for life in the water. Sound travels farther and 4 1/2 times faster in water than in air, is little affected by murkiness or depth, is an excellent means of echolocating objects and an excellent medium for socialization. Thus it seems quite likely that very elaborate sound processing is a major function of the large cetacean brain. A large part of the human brain is involved in processing visual information. Light waves vary in length from about 4 to 8 hundred thousandths of a centimeter, but sound waves used by cetaceans vary from about a centimeter to over 70 meters. Perhaps the relative physical grossness of sound would require more neurons and cell assemblies in order to process sound data with a resolution of detail approaching that of vision.
Whatever intelligence cetaceans may have appears to be different from ours. This is indicated by the ease with which they are killed by whalers or trapped in fishing nets. Yet the size and complexity of their brains indicate enormous activity and possibly an extraordinary rich mental life. Their mental life is probably much more involved with social relationships than problem solving. The intelligence of dolphins is indicated by their love of play, complex social behavior, and the ease with which they learn tricks in captivity. Gregory Bateson1has shown that dolphins not only can learn new tricks, but also can learn that they are supposed to learn a new trick in new sessions. The new trick may even be the negative of behavior which was rewarded earlier. He notes that the dolphin cannot do this by conditioning from a single sample, but must do it from the structure of a complex sample. This kind of learning operates on a higher logical level than learning from the repetition of a type of instance.
Humpback whales sing complicated songs that are as long as half an hour. The songs evolve from year to year, indicating an impressive memory. The whales do not sing in their summer feeding grounds, but they begin the previous season's songs when they return to their winter grounds and gradually change them as the season progresses. They are able to memorize all the complicated sounds of a song, as well as new modification, for at least six months as a basis for further improvisations.
|
|
The cetacean brain has evolved in a simpler, less kinesthetically manipulable, and less demanding environment than that in which the human brain arose. This contrast is reflected in the different organizations of the two brains. The lamination and regional differentiation of the cerebral cortex are comparably developed in both species, although differently arranged. In cetaceans the areas of the cortex receiving information from sense organs, especially those representing the ears and face, are very large. Motor control occupies less cortical area than in the human brain, indicating orientation more toward perception and less toward action and motor skills than in humans.
Each hemisphere of the human brain has three basic, concentric formations representing successive stages of neural evolution. The innermost and oldest, the rhinic lobe, supports the basic survival functions. The middle, or limbic lobe, is the seat of emotional activity and motivation. The third and largest, the supralimbic lobe, is the seat of rational processes. Cetaceans have a fourth formation, the paralimbic lobe, between the limbic and supralimbic lobes. This is a unique, very large feature not found in other mammals, a specialized region in which all sensory and motor areas are represented together. In the human brain the different sensory areas are widely separated. For example, the motor area lies next to the touch area but is separated from all other sensory regions. An integrated perception of sight, sound, and touch is dependent on electrical impulses in the neurons that travel long distances in which time and information are lost. Apparently, the paralimbic lobe supports the very rapid formation of integrated perceptions.
It has been suggested that the scattering of sense projection areas in the human brain evolved from the need for quick danger response. Each of the sensory areas is bordered by interpretation areas which deal with only one sense. When a sound suggesting danger is registered, the neural system signals alarm before receiving visual conformation. Neither does vision wait for sound or touch. Our intellectual functions are based on these separate sense interpretation areas. Perhaps this favors abstract thought in the human mind, while the paralimbic lobe favors an immensely rich apperception. If the cetacean brain has fewer separate association areas, the cetacean mind may have a smaller variety of concepts. It would be more difficult for whales to have abstract ideas because there would be less variety of basically different things to compare. Hence they would tend to have less problem solving ability and less intelligence than humans, although not less, and possibly greater, cerebral activity.
|
||||||||||
| Figures 3.A - 3.E. Cut-away images of bottlenose dolphin brain structure. | ||||||||||
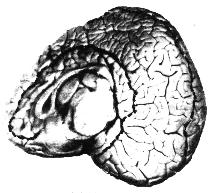


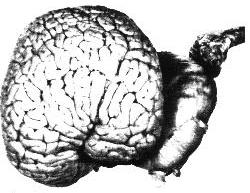
Sterling Bunnel, a psychiatrist, suggested that the evolution of the large cetacean brain originated in the needs of river dolphins to find their way about by echolocation in the murky waters of rivers and estuaries.2The Ganges River dolphin is blind. In rainy seasons it leaves the river, pursuing fish through flooded forests in very turbid water. It is necessary in these rangings to form detailed maps to prevent being trapped by receding waters. Thus the auditory and tactile senses and the general association areas of the brain in which memories are stored have become highly developed. It is true that small-brained bats have good echolocation for capturing insects and for navigating in caves, but sonic memory maps are not needed to capture insects and bats spend only a few minutes each day navigating to and from their fixed roosts. The very early whales, which evolved from land animals that adapted to the sea, had small brains. If large-brained whales evolved in the rivers and estuaries, they could have returned to the sea and replaced the more primitive whales. In the sea the cetacean brain entered a second stage of evolution and continued to increase in size. The continuing increase may have been a result of social selection, because the demands of survival in the marine environment do not seem sufficient to evolve such huge brains. Elephants have a complex social life, and their brains may have evolved through social selection after their great size made their lives more free of danger. Our own large brains may have evolved not only from the advantages of problem solving ability and deceptive behavior, but also from social selection of desirable psychic qualities such as empathy and sensitivity. Highly developed social interaction, including deception, requires the ability to image, model, and remember the psychic state of other beings, and such ability requires a very large amount of neural activity.
Bunnell3believes that the cetacean's senses and great memory capacity, developed for navigation in murky water, are ideally pre-adapted for social communication. The echolocation apparatus not only can project complex signals, but has the capability for detailed three-dimensional perception of the physical and emotional state of another cetacean's entire body. Echoes arise from the interior body as well as the skin of another cetacean. Actually, stronger echoes arise from the air-filled cavities and the bone of an animal than from the skin. This enables the pulsations of the vessels and viscera, and hence the emotional state and health of an animal, to be easily perceived, adding a new dimension of social interaction.
Thus the need for elaborate sound processing is the most likely reason for the great development of the cetacean brain. Sound is used very extensively by cetaceans, and water is much more pervaded by sound than light. Sam Ridgway, a dolphin researcher at the Naval Ocean Systems Center in San Diego, believes that much of the great development of the dolphin auditory system is a result of a need for great precision and speed in processing sound.4But surely dolphins are also processing more sound data and processing them far more elaborately than bats or owls. Indeed, if sound data are to be processed into sensory field comparable to vision, the data would need to be processed very rapidly and precisely. Let us consider in more detail some of the possible uses of elaborate sound processing.
The two basic possible uses for elaborate sound processing are communication and perception. The whale's environment does not seem sufficiently complex to require a vast amount of information communication. But the possibilities for social communication and selection of feelings, emotions, and aesthetic sound patterns are extremely rich. It seems likely that they frolic with their minds as well as with their bodies. Perhaps they can create sound so elaborate, beautiful, and satisfying that they have selected that capability just as birds of paradise have selected spectacular feathers. Such feathers have no survival advantage and no severe disadvantage in a benign environment, but they are strongly preferred by the birds. There are occasions when animals do act for immediate satisfactions. This is vividly demonstrated by the extreme case of self-stimulation, to exhaustion, by rats which are able to activate electrodes implanted in the pleasure centers of their brains. When an action is not automatic as breathing, reflex, or nest building by insects, it may be done for pleasure or avoidance of pain rather than for some direct survival advantage. The survival purpose of an action may arise secondarily rather than by choice. When young animals play, they are not aware that they are developing social and survival skills. Elaborate behavior or effects may be selected for gratification when there is no definite survival disadvantage.
It is possible that whales use advanced sound forms and are involved in aesthetics that are inconceivable to us. The functions of the paralimbic lobe may have the effect of fusing the various senses in the manner that hearing speech fuses sound and meaning. The areas of the human brain that process sound and meaning lie close together. We do not hear a word and then translate it unless we are struggling with a new language. We hear meaning. Thus the cetacean's senses may be fused by the paralimbic lobe to produce a direct apperception of a richness and beauty unknown to us.
Bunnell5notes that vision is a special sense which gives simultaneous information in the form of pictures with poor time discrimination, and that our auditory sense has very limited space perception but good time discrimination. Thus our languages are composed of sounds in temporal sequences. He suggests that the cetacean auditory system is essentially spatial with much simultaneous information. Cetacean communication apparently consists of complex sounds perceived as a unit. These communications are probably not a syntactical language of technical information or abstract concepts. Perhaps they project ideograms representing entire social situations, elaborate emotions, extensive sequences of events presented as a picture, or all of these in one extraordinary unit. The paralimbic lobe may well have enormous memory capacity, which would expand potential size and complexity of an ideographic vocabulary. Bunnel further speculates that the cetacean echolocation system produces detailed images of objects, and that perhaps they can recreate these sounds and directly project sonic images to one another.
Projection of images of one's bodily state, past, present, and future, and of the states of others would be a powerful way of expressing feelings and emotions. Cetacean language may be somewhat similar to Chinese ideograms in which pictures are equated to ideas. It may be possible to confirm some of these conjectures by comparing audiograms of reflected and projected sounds. Similar patterns in the sounds would indicate communication of perceptions, especially if the patterns were very complex. The social, emotional and aesthetic life of humans would be greatly enriched by the ability to project visual images to others that would be as detailed as the actual viewing of an object, scene, or situation.
The fusing of received and projected sonic messages with other sensory experiences has vast possibilities for aesthetic and social experience. Some idea of what this may be like is suggested by the special experience of eye contact. They are special because quick, subtle movements and light changes interact within the actual time of the mental processes. Such real time interplay is much diminished in communication through delayed serial clusters of speech. The intimate interactions of eye contact seem to fuse with the emotions of the moment. The nature of sound allows perception from all directions and makes it possible for more than two cetaceans to enter into such communication. If three or more people could engage in eye contact at the same time, could generate and project meaningful patterns of light both bodily and ideographically, all in real time and fused as intimately as meaning and sound are fused in speech, then humans might have experiences similar to the possibilities of cetacean social life.
The other major use for elaborate sound processing is location and perception of objects. The sperm whale has the largest brain of all and it is also the only whale which dives to great depths for prey, at least to 3600 feet and possibly to two miles. This is certainly an environment in which sonic echolocation and very detailed sophisticated sonic perception would be of prime usefulness, especially since the whale can remain at such great depths for less than 45 minutes. The obvious usefulness of sonic perception at great depths is consistent with the sperm whale's great increase in brain size. This may be a third step in the evolution of the cetacean brain. The enormous neural development for social life, which may have arisen after river dolphins returned to sea, would be a natural basis for further evolution needed for deep diving.
Our basic visual field is two-dimensional, which can be appreciated by looking at bright light with closed eyes. The third dimension of visual space is given by a system of distance values assigned through elaborate cues, such as ocular convergence, binocular disparity, texture density gradients, motion parallax, superposition, and illumination direction. Another three-dimensional sensory space is the kinesthetic space in which is experienced when moving about in a dark room. This space is much more highly developed in blind people. We have a poorly developed sonic space because the sound elements have very little connectivity, although a continuous, rapidly varying pitch can give a suggestive glimmer of a one-dimensional sonic space. Blind people are able to make remarkable use of sound for gauging their surroundings, but this is not a sonic space. It is rather a kinesthetic space closely associated with sonic data. Sensory spaces are not physical space, although they are coordinated with it. Visual space can be distorted by lenses, drugs, or brain injury, but physical space is distorted only by extreme conditions, such as very powerful gravitational fields, which are not encountered in ordinary life.
It seems possible that, if cetaceans can assign definite directions to sounds, they can convert sound stimuli into neural patterns that generate a three-dimensional sonic sensory space. If this is true, then the whale would experience itself immersed in a three-dimensional sonic space similar to our immersion in three-dimensional. That is, it would "see" with sound. The immediate sonic field would probably surround the whale, unlike our immediate visual field which is in front of us, although the greatest clarity and acuity would be in front because the head region is better innervated for sound reception. The head is also the source of sound projection, and the echoes of projected sound are more unidirectional in the high frequencies.
Objects would be located in this space as we locate objects in visual space. There would be both sources and reflectors of sound, just as there are emitters and reflectors of light in the visual world. If sound has sufficiently high frequency, it can be reflected with such coherence that an object, e. g., a fish, would be perceived as a fish in a manner comparable to our seeing a fish. The selective reflectivity of different frequencies by various kinds of objects might even be perceived as qualities analogous to color.
The cetacean's sonic space would differ structurally from human visual space. Perception in sonic space would have less acuity than in visual space and the acuity would vary with the frequency of the sound. The distance range of detectable sound in water varies from a few feet for extremely high frequencies to many miles for very low frequencies. There is a noticeable time delay of one second per 5000 feet in the transmission of sound in water. The frequency of sound from a source is increased or decreased by the Doppler effect when the source moves toward or away from the observer. In contrast, perception in visual space is much sharper, has practically the same acuity and range for all wavelengths of visible light, and has no time delay or Doppler effect in ordinary life. These differences between sonic and visual spaces are evident because sound is far slower than light and has greater wavelengths. An object cannot reflect a wave whose length is greater than the size of the object. Dolphins can emit and detect sounds at least as high as 120,000 cycles per second. Sound at that pitch can be used to detect objects as small as an inch. This is a resolution of about 7 minutes of arc at 20 feet, although proportionally less at closer distances. Dolphins can also determine the direction at this sound with about the same accuracy. It is possible that the dolphin can use even higher frequencies which would give still better resolution. The resolution of human vision is about a minute of arc. It is also quite possible that the dolphin combines its sonic and visual spaces into a single sensory space which would have more structural uniformity and better resolution at close ranges. Combining these spaces may be one of the functions of the paralimbic lobe.
Far more must be known about the mechanism for generating visual space from the neural activity of the visual cortex before we can understand the manner, or even the possibility, of generating a sonic space. Even then we may know if the quality of the space is sonic or visual, i. e., whether it would be occupied by sound or by visual sensa supported by sonic input. All mental activity is based on electrical impulses in nerve cells, and all types of these cells have the same kind of impulse. The sensory quality of an experience depends on the processing and organization of the impulses. If the nerves of the inner ear were connected to the visual cortex, sounds would be experienced with a visual quality.
Cetaceans do not have an organ concentrating an array of sound detectors similar to the array of rods and cones in the retina. Dolphins have two inner ears that receive sound via fatty deposits inside the jawbone and a region of the forehead called the melon that possibly functions as a sonic focusing device. If they experience a sonic space of two or more dimensions by means of this apparatus, that space must be supported by some means of storing the data as they are received through the two ears. Somewhat analogously, the visual impressions from the moving spot of light that scans the surface of a television tube is held by the observer through persistence of vision for about 1/10 of a second, long enough for the scan to cover the entire tube and enable the observer to experience an image. If this were not so, we could perceive only a moving spot. The dolphin's two inner ears cannot construct a two-dimensional field from a fixed position, but head movement, swimming, and deliberate beaming of emitted sound in different directions could generate variations sufficient to support such a field. These movements would also widen the perceptual field for the high frequencies. It is also possible that cetaceans can detect sound through pressure variations on the skin. If this is the case, it might be possible for them to have a two-dimensional space of sensations via an array of sensors extending over the body surface. Such an array has the advantage of greater sonic data input for a given time interval, and data based on the array is more easily organized into a sensory space of more than one dimension.
|

The field of vision is experienced as a coherent continuum. This seemingly simple property cannot arise from a mere array of sensory elements alone. There must be some process which binds the elements, or they would be independent and would not appear together as a single expanse. This coherent field of vision may be supported by some sort of neural scanning mechanism. This is suggested by an interesting visual effect which can be induced by a flashing stroboscopic light. When an observer looks at such a light flashing between 6 and 30 times per second, his visual field becomes filled with complex, changing patterns such as dark or colored zigzag lines vibrating on a light background. Other patterns may be like the spokes of a wheel or a whirlpool. The zigzag lines may sweep from one side of the visual field to the other or form intricate mosaics. These effects do not come from the retina. This was demonstrated by Gray Walter,6a physiologist, who found that the electrical pulsations in the retina show no unusual effects under such flashing stroboscopic lights. Similar phenomena occur on a television screen when a studio illuminated by a flashing light is monitored by a television camera. Complex patterns of dots and lines dart and zigzag across the screen. This happens because the moving electron beam is scanning the screen to form the picture can illuminate the screen only during the very short intervals when the light is on in the studio. Thus only disconnected bits of the beam's rapid path over the screen are visible. The particular form of the patterns depend on the particular nature of the scanning mechanism. Possibly the whirling spiral that is often seen under stroboscopic flicker indicates the type of scan in the neural system which supports the visual field. This scan is not, of course, anything like a moving electron beam. It would perhaps be some sort of rhythmical firing of banks of nerve cells.
If an organism has a two-dimensional sensory field supported by a neural scanning mechanism, such a mechanism might be detected by electrodes in an appropriate part of the brain which can register the scan's electronic signature. If the firing of banks of cells in the visual cortex of a mammal has some characteristic erratic signature under stroboscopic light flashing at critical frequency, such a signature may also characterize other two-dimensional sensory fields and could be used to test for their existence. If this is the case and a dolphin were subjected to extremely short bursts of sound at constant critical intervals, and erratic firing of banks of auditory cells, similar to that of visual cells under stroboscopic flicker, might become evident. If the sonic data under the scan are stored longer than the duration of a single complete scan, then the sonic field could be two-dimensional with elaboration to three dimensions through cues such as the Doppler effect, modification of sound reflections by the shape and texture of surfaces, and by differential time delays of reflection from objects at various distances.
Suppose a two-dimensional space of sonic data with its three-dimensional cues can be held in storage for a few seconds while other, similar three-dimensional spaces are rapidly and continuously constructed and stored. The fields might be stored by repeated firing of banks of cells. Suppose further that a higher scan surveys this continuum of three-dimensional spaces and integrates them into a single manifold. This would have the effect of a four-dimensional sensory space in which span of time would have a spatial representation. Such a feat would require and enormous neural system. This conjecture is, of course, very speculative, but here may be some natural base for such a development. The huge supralimbic and paralimbic lobes of cetaceans very likely have enormous memory capacity, and if they store data to support sonic fields, this could be a base for a next stage of perception. If cetaceans do experience three-dimensional sonic spaces, then some survival or social advantage for another dimension of sonic space might induce such a development. The sperm whale's largest brain size suggests the greatest development for processing sonic data. If there is any survival advantage to the ability to experience four-dimensional sonic space, it may well be associated with deep diving.
It is not certain that this ability would be of any technical advantage to the whale. The sperm whale's limited time at great depths must be spent very efficiently to capture enough prey to support so large a creature. A capacity for four-dimensional sonic perception would allow it to perceive the trajectories of its prey imaging dynamic processes that would enable it to capture prey more effectively. Possibly a whale would also be able to monitor the activities of other prey while it is actually feeding. Such perception has an advantage at great depths because there is no continuous field of contiguous objects such as we experience in our daily life, but separate objects moving about in a sonically dark, or relatively dark, background. A four-dimensional sensory space would be most effective if its span represented a few seconds. A longer span would clutter perception. A whale's aesthetic and social life would be enormously enhanced by a four-dimensional sonic space. Perhaps they could create music, sonic arabesques and sculpture, or social messages beyond our comprehension.
Humans experience a slight, crude four-dimensional visual space. This is manifest when we see the trajectory of a rapidly moving light in a dark room. We would be unable to see a picture on a television screen if our perceptions were strictly from instant to instant. We would see only a series of dots as the electron beam scanned the tube. The slight persistence of hearing combined with the running memory enables us to experience rhythms. Thus we operate in a crude four-dimensional space, but its span represents only 1/10 of a second.
If tests indicate that cetaceans experience sonic fields, it may be possible to make tests indicating four-dimensional sonic perception. This might be done by implanting electrodes in the auditory region of a cetacean's brain and presenting objects for it to echolocate which arouse a definite electronic signature. The rapid repetition of the signature for a sustained time would suggest a four-dimensional sonic perception. That is, we might try to determine of a sound pattern "reverberates" in the cetacean's brain. In another possibility, a signature might be linked to a very transient event, such as an abrupt change of direction by a darting squid, and the time span of the event found to be less than that of the signature repetition.
Cetaceans may be less proficient problem solvers than humans, but they surely have enormous cerebral activity. A creature is a being when it experiences a conscious sensory field, and is a person when it has a concept of the self and an awareness of its own self. The self may have arisen from the needs for survival planning by problem solving creatures in dangerous environments. If this is true, then cetaceans are less likely that humanoids to have selves. But the sense of self would enrich cetacean life and would be very useful in social competition. If cetaceans' sonic transmissions contain some symbol for the sender and for each individual, i. E., a name, personhood would be suggested. All cetaceans are almost certainly beings. Perhaps some are prodigious beings, and conceivably persons.
Boulder, Colorado (1992)
|
1 "Observations of Cetacean Community," Gregory Bateson in Mind in the Waters. Assembled by John McIntyer. Scribners/Sierra Club, 1974.
2 "The Evolution of Cetacean Intelligence," Sterling Bunnell in Mind in the Waters. Assembled by John McIntyer. Scribners/Sierra Club, 1974.
3 Op. Cit., "The Evolution of Cetacean Intelligence."
4 "Physiological Observations on Dolphin Brains." Sam H. Ridgway in Dolphin Cognition and Behavior: A Comparative Approach. R. Schusterman, J. Thomas, and F. Wood (Eds.). Lawrence Erlaum associates, Hillsdale, new Jersey, 1986.
5 Op. Cit., "The Evolution of Cetacean Intelligence."
6 The Living Brain. Gray Walter. P. 102. Norton, 1953.











No comments:
Post a Comment